
Abiotic Stress Signaling and Responses in Plants. As sessile organisms, plants must cope with abiotic stress such as soil salinity, drought, and extreme temperatures. Core stress-signaling pathways involve protein kinases related to the yeast SNF1 and mammalian AMPK, suggesting that stress signaling in plants evolved from energy blogger.com by: 11/17/ · From this review, we have constructed a simplified working model summarizing some of the known plant signaling responses to abiotic stress (Figure 2). Much of the signaling involves phosphorylation cascades that react quickly in the plant cell, emphasizing the need for proteomics data as well as transcriptomics data in future blogger.com by: Drought and salinity are the abiotic stresses most studied in quinoa; however, studies of other important stress factors, such as heat, cold, heavy metals, and UV-B light irradiance, are severely limited. In the last few decades, the incidence of abiotic stress has been accentuated by the increase in unpredictable weather blogger.com by: 59
Effects of abiotic stress on plants: a systems biology perspective | BMC Plant Biology | Full Text
Abiotic stresses, such as low or high temperature, deficient or excessive water, high salinity, heavy metals, and ultraviolet radiation, are hostile to plant growth and development, leading to great crop yield penalty worldwide.
It is getting imperative to equip crops with multistress tolerance to relieve the pressure of environmental changes and to meet the demand of population growth, as different abiotic stresses usually arise together in the field.
The feasibility is raised as land plants actually have established more generalized defenses against abiotic stresses, including the cuticle outside plants, together with unsaturated fatty acids, reactive species scavengers, molecular chaperones, and compatible solutes inside cells. In stress response, they are orchestrated by a complex regulatory network involving upstream signaling molecules including stress hormones, reactive oxygen species, gasotransmitters, polyamines, phytochromes, and calcium, as well as downstream gene regulation factors, particularly transcription factors.
Land plants are living in an inherently harsh environment ever since their emergence. A large variety of physical or chemical factors are hostile to them, including low or high temperature, deficient or excessive water, high salinity, heavy metals, and ultraviolet UV radiation, among others.
These stresses, collectively referred to as abiotic stresses, are posing a severe threat to agriculture and the ecosystem, accounting for great crop yield loss Wang et al.
Salt stress is the most stubborn one magnified by ever-increasing salinization of arable land worldwide Munns and Tester, ; Yuan et al. Most plants cannot survive when NaCl concentrations exceed mM Flowers and Colmer, ; Zhou J.
et al. More seriously, it is interlinked with drought, another global issue, which can be aggravated by extreme temperatures Ashraf and Foolad, ; Slama et al. Due to their sessile nature, plants have to confront the stresses and develop potent adaptive tactics to avoid or tolerate their adverse effects so as to survive and to thrive. Plenty of cellular, physiological and morphological defenses have been established. The most apparent one is the cuticle, a universal outmost shield Shepherd and Wynne Griffiths, ; Yeats and Rose, ; Fich et al.
It is also impressive that recretohalophytes even evolved a specialized organ to excrete salt, as represented by the epidermal salt gland of Limonium bicolor Yuan et al. Tremendous progress has been made toward understanding the biochemical and molecular mechanisms underpinning the defenses, owing to forward and reverse genetic approaches as well as genome-wide analyses conducted on various model species review article on abiotic stress in plants the classical model Arabidopsis thaliana and its extremophyte relative Thellungiella salsuginea that has exceptional multistress resistance Amtmann, ; Wang J.
It is thus emerging that desaturation of membrane lipids, activation of reactive species RS scavengers, induction of molecular chaperones, and accumulation of compatible solutes are more generalized and conserved cellular defense responses. This is in line with the fact that membrane injury, RS damage, protein denaturation, and osmotic stress primarily dehydration can be provoked by a multitude of abiotic stresses.
In stress response, these defenses are orchestrated by a complex regulatory network involving upstream signaling molecules including stress hormones [e. Figure 1. The general defense systems and the underlying regulatory network in botanic responses to abiotic stresses.
Different abiotic stresses, such as cold, heat, drought, flood, and salt can provoke common cellular disorder and secondary stresses, review article on abiotic stress in plants, including membrane injury, reactive species RS damage, protein denaturation, and osmotic stress, which are also interconnected with each other. Accordingly, review article on abiotic stress in plants, land plants have resorted to unsaturated fatty acids, RS scavengers, molecular chaperones, and compatible solutes.
Some compatible solutes may also be involved in counteracting other adverse effects, as indicated with dotted inhibitory lines.
Besides, the cuticle serves as the universal outermost shield. Notably, phytochrome B PHYB is emerging as a negative regulator in stress tolerance. In the field, plants are routinely review article on abiotic stress in plants to an unpredictable combination of different stresses rather than a single one Slama et al. Particularly, on demand of population growth, it is getting imperative to equip crops with multistress tolerance. In this section, five general botanic defenses against abiotic stresses will be addressed, comprising cuticle as outermost shield, unsaturated fatty acids UFAs as membrane modulator and review article on abiotic stress in plants precursor, RS scavengers that govern RS homeostasis, molecular chaperones that stabilize proteins and subcellular structures e.
Chances to acquire multistress tolerance based on them are given in Table 1. Notably, cuticular waxes, UFAs, antioxidant compounds, and compatible solutes are review article on abiotic stress in plants economic traits as well. For example, waxes are raw materials of manifold products including biofuels, cosmetics, detergents, plastics, and pharmaceuticals Lee and Suh, Therefore, increasing their yield actually serves a double purpose in crop improvement.
Table 1. Chances to generate multistress tolerance based on the general defenses. Land plants have an exterior translucent lipid structure, namely the cuticle, sealing the aerial surfaces of their organs. The thin hydrophobic layer is basically a cutin matrix filled with and coated by cuticular waxes.
As the primary interface between plant and environment, the cuticle plays critical roles in restricting liquid and gas fluxes, defending pathogen and insect attacks, and resisting various abiotic stresses.
It is an elegant innovation of land plants to deploy an review article on abiotic stress in plants shield derived from simple molecules, which is fundamental to their success in terrestrial colonization for review, see Shepherd and Wynne Griffiths, ; Yeats and Rose, ; Fich et al.
By contrast, cell wall, the second barrier that is actively remodeled under abiotic stresses Shen et al, review article on abiotic stress in plants. The cuticle is exclusively created by epidermal cells. Typically, cutin is a macromolecular polyester of C16 or C18 oxygenated fatty acids FAswhereas waxes are a complex mixture of C24 to C34 FA derivatives, including alcohols, aldehydes, alkanes, esters, and ketones. Their biosynthetic pathways are nearly resolved and have been well documented see Pollard et al.
Notably, review article on abiotic stress in plants, two distinct modification pathways are involved in wax generation, the alcohol-forming or acyl-reduction pathway for primary alcohols and esters, together with the alkane-forming or decarbonylation pathway for aldehydes, alkanes, secondary alcohols, and ketones. To assemble the apoplastic cuticle, these materials need to be exported from the ER to the plasma membrane PMand then across the PM through the cell wall onto the outer surface where cutin monomers polymerize and wax members crystallize.
Membrane vesicle trafficking McFarlane et al. The likely extracellular relays for traversing the hydrophilic cell wall are non-specific lipid transfer proteins nsLTPsa group of small and basic proteins bearing a hydrophobic pocket for lipid binding. Indeed, two glycosylphosphatidylinositol-anchored LTPs, LTPG1 and LTPG2, as well as a secreted one, TsnsLTP4 from Thellungiellahave been reported to be implicated in wax deposition Debono et al.
Then comes the last procedure of cutin production, i. The crosslink is formed directly or via a bridging molecule, e.
However, both the polymerization mechanism and the polyester architecture have been longstanding enigmas. Nevertheless, since the fruit of CUS1 null mutant, cutin deficient 1 cd1are not fully deprived of cutin, non-enzymatic mechanisms cannot be ruled out yet Yeats et al. Drought tolerance is closely associated with wax accumulation in a wide variety of plant species see reviews Borisjuk et al. With respect to multistress tolerance, however, wax composition makes a difference and the alcohol-forming pathway seems to outperform the alkane-forming one.
It is highly possible that in CER1-overexpressing plants, cold tolerance was compromised, in parallel with pathogen defense and leaf growth, although water deficit resistance was improved Bourdenx et al. Indeed, increased level of n -alkane coupled with decreased level of primary alcohols led to cold susceptibility and growth retardation.
In contrast, higher contents of both resulted in better viability under drought and freezing without disturbing plant growth Zhang et al. Besides, fatty acyl-CoA reductases FARs that produce primary alcohols could be up-regulated by cold, heat, polyethylene glycol PEG; dehydrationABA, methyl jasmonate MeJAand fungal infection in wheat Triticum aestivum Chai et al.
The stress-resistant performance of FAR-overexpressing plants is thus intriguing. Another good candidate for genetic engineering might be CER6, the major 3-ketoacyl-CoA synthase KCS that catalyzes the initial and rate-limiting condensation step of FA elongation, as its overexpression could elevate total wax output with little alteration of the composition.
Of note, it was not the cauliflower mosaic virus CaMV 35S promoter but the native one review article on abiotic stress in plants could drive CER6 expression high enough to achieve significantly greater wax quantity in transgenic Arabidopsis Hooker et al. In addition to the enzymes, transporters can also be taken into consideration.
Actually, TsnsLTP4responsive to cold, heat, NaCl, PEG, and ABA, has been introduced into Arabidopsis and augmented its tolerance to drought and salt Sun et al.
Notably, more chances reside in manipulating the TFs that control cuticle generation, which will be discussed in the end. FAs thereby have a profound impact on membrane properties. Particularly, their unsaturation degree is a major determinant of membrane fluidity in that UFA chain will create a kink at a cis -double bond, which serves as steric hindrance in intermolecular package leading to a more fluid state Hazel, ; Mikami and Murata, Membrane fluidity is susceptible to various abiotic stresses, extreme temperatures in particular.
Both cold-driven rigidification and heat-driven fluidization can cause biomembrane dysfunction, as exemplified by protein deactivation and ion leakage Hazel, Cytoskeleton destabilization is also a direct consequence Sangwan et al. Membrane remodeling is thus of especial importance in plants, which are poikilothermic organisms, review article on abiotic stress in plants. Indeed, adjusting the unsaturation degree of the FA tails in bilayer interior is favored by plants in offsetting thermal perturbations to maintain the optimal range of fluidity.
Particularly, there is a very close relationship between chilling tolerance and the unsaturation level of chloroplastic phosphatidylglycerol PG for review, see Nishida and Murata, ; Iba, In thylakoid membranes that are biased toward glycolipids, PG is the only phospholipid species present. Actually, it is an indispensable component of the membrane-bound photosynthetic apparatus including Photosystem II PSII Wada and Murata, PSII is vulnerable to photoinhibition, in which the D1 protein of the reaction center is bound to continuous photodamage followed by repair via proteolysis and synthesis Takahashi and Murata, ; Liu et al.
Desaturation of PG has been shown to protect PSII against cold-enhanced photoinhibition, which contributes to chilling tolerance Moon et al. This is also applicable to other stresses that can intensify photoinhibition Takahashi and Murata, Upon NaCl treatment, for instance, alleviated photoinhibition of PSII pertained to increased contents of UFAs in membrane lipids including PG Sui et al.
Indeed, specifically elevating the unsaturation level of PG accelerated the turnover of the D1 protein Sun et al. It is noteworthy that polyunsaturated UFAs, upon liberation by lipase from glycerolipids, also serve as the raw material of oxylipins, bioactive molecules involved in diverse physiological processes, including stress resistance see review Savchenko et al. Particularly, linolenic acid gives birth to jasmonic acid and its derivatives, namely jasmonates JAsa group of stress hormones with a well-understood role in launching wound response.
There is emerging evidence that JA is also implicated in defense against other stresses, such as salt Ryu and Cho, ; Yang et al. It is of economic interest that applying MeJA to fruits and vegetables can reduce chilling injury, which is conducive to the maintenance of their post-harvest quality González-Aguilar et al.
Unsaturation is administered by position-specific FA desaturases FADs. To synthesize C18 UFAs that are more active in stress tolerance, C18 product of de novo FA synthesis linked to acyl-carrier protein ACPnamely ACP, is first converted to 9 -ACP by stearoyl-ACP desaturase SAD. After being incorporated into glycerolipids, 9 is processed to 9, 12 by ω-6 desaturases and then to 9, 12, review article on abiotic stress in plants, 15 by ω-3 desaturases Murata and Los, To resist various stresses like cold and wounding, the level of is usually elevated.
This is much achieved by activation of ω-3 desaturases comprising ER-associated FAD3 and plastid-localized FAD7 and FAD8, as revealed by mounting evidence Shi Y.
On the contrary, review article on abiotic stress in plants, gene silencing of FAD7 enabled transgenic tobacco Nicotiana tabacum to abide high temperatures Murakami et al. Hence, inducible overexpression of FADs might be better so that heat tolerance can be covered. The unsaturation level of chloroplast PG is otherwise determined by the substrate specificity of plastid glycerolphosphate acyltransferase GPATwhich catalyzes the first reaction to esterify FAs into glycerolipids.
A preference for ACP matters as the second reaction always utilizes ACP Nishida and Murata, ; Iba, Interestingly, GPATs from Arabidopsis resistant and squash Cucurbita moschata sensitive respectively assimilated the chilling behavior of tobacco intermediate Murata et al.
Actually, under saline situations, S. salsa also exhibited chilling resistance Cheng et al. Moreover, ACPs, as essential cofactors for FA synthase FASSAD and GPAT, are also implicated in the alteration of FA composition. Transformation with AhACP1 from peanut Arachis hypogaea into tobacco resulted in significantly higher contents of and accompanied with more tolerance against cold Tang et al. The two types of RS are intertwined with each other.
RCS can arise from ROS-induced lipid peroxidation, while ROS can be raised by RCS activities the other way round.
plant respond to biotic and abiotic stress
, time: 2:05Plant signaling in biotic and abiotic stress | Journal of Experimental Botany | Oxford Academic
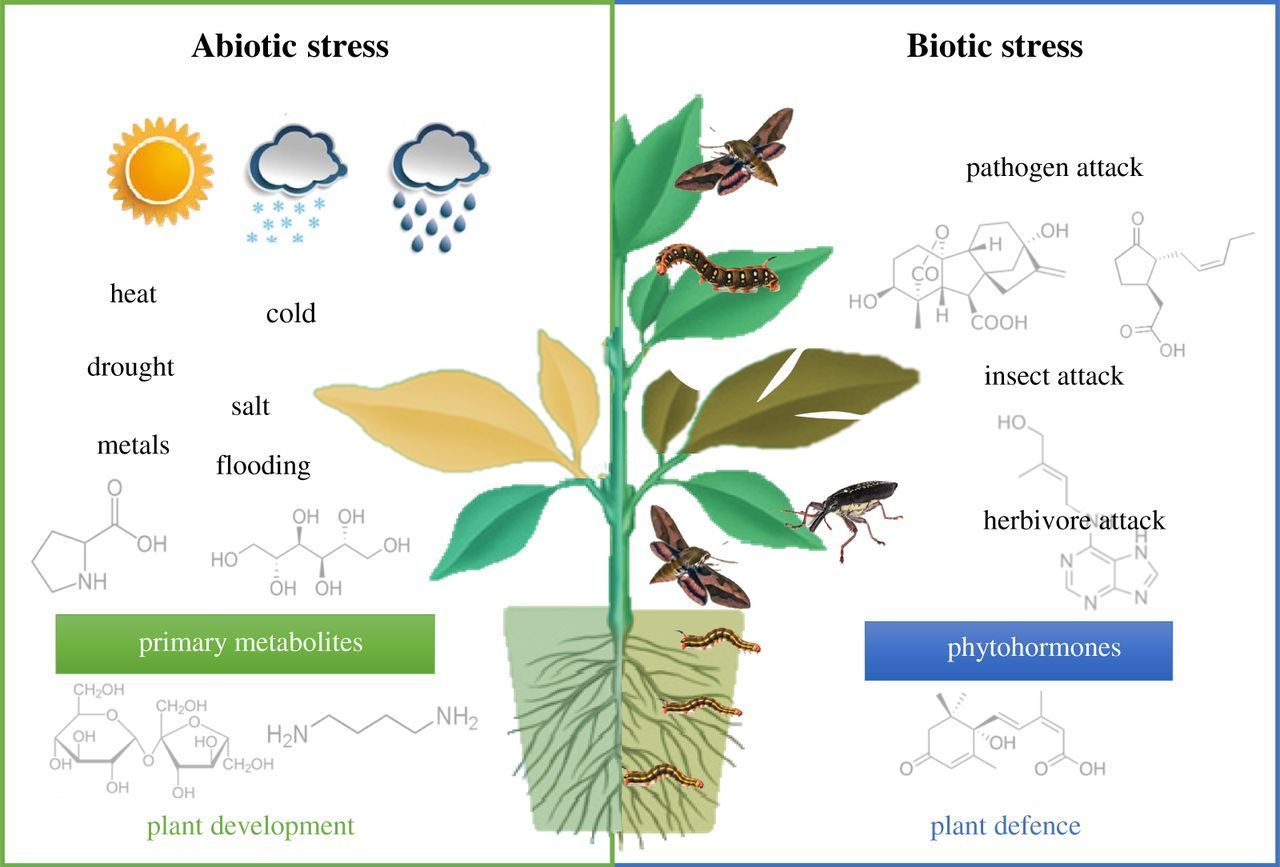
3/12/ · Abiotic and/or biotic stress combinations (i.e. the study of how plants respond to two or more different environmental stressors that impact them simultaneously) is a rapidly developing field in plant biology (highlighted in this issue by Zandalinas et al., ). It is perhaps one of the best examples for the concept of signal transduction blogger.com by: 2 Abiotic Stress Signaling and Responses in Plants. As sessile organisms, plants must cope with abiotic stress such as soil salinity, drought, and extreme temperatures. Core stress-signaling pathways involve protein kinases related to the yeast SNF1 and mammalian AMPK, suggesting that stress signaling in plants evolved from energy blogger.com by: Ozone (O3) is a major abiotic stress which severely affects the growth and development of plants. In order to cope up with ozone stress, plants exhibit a plethora of morphological, physiological




No comments:
Post a Comment